Goditi lo spettacolo emozionante delle slot online con jackpot impressionanti su siti scommesse non aams, dove sicurezza e affidabilità sono al primo posto per un'esperienza di gi.
Scopri l'emozione di giocare con veri croupier grazie a Lolajack, dove l'atmosfera del casinò in tempo reale trasforma ogni partita in un'esperienza unica e coinvolgente.
godere dell’acqua arricchita di idrogeno direttamente a casa tua, migliorando la tua energia,
riducendo lo stress ossidativo e supportando uno stile di vita attivo e sano.
È semplice, efficace e trasformativo.
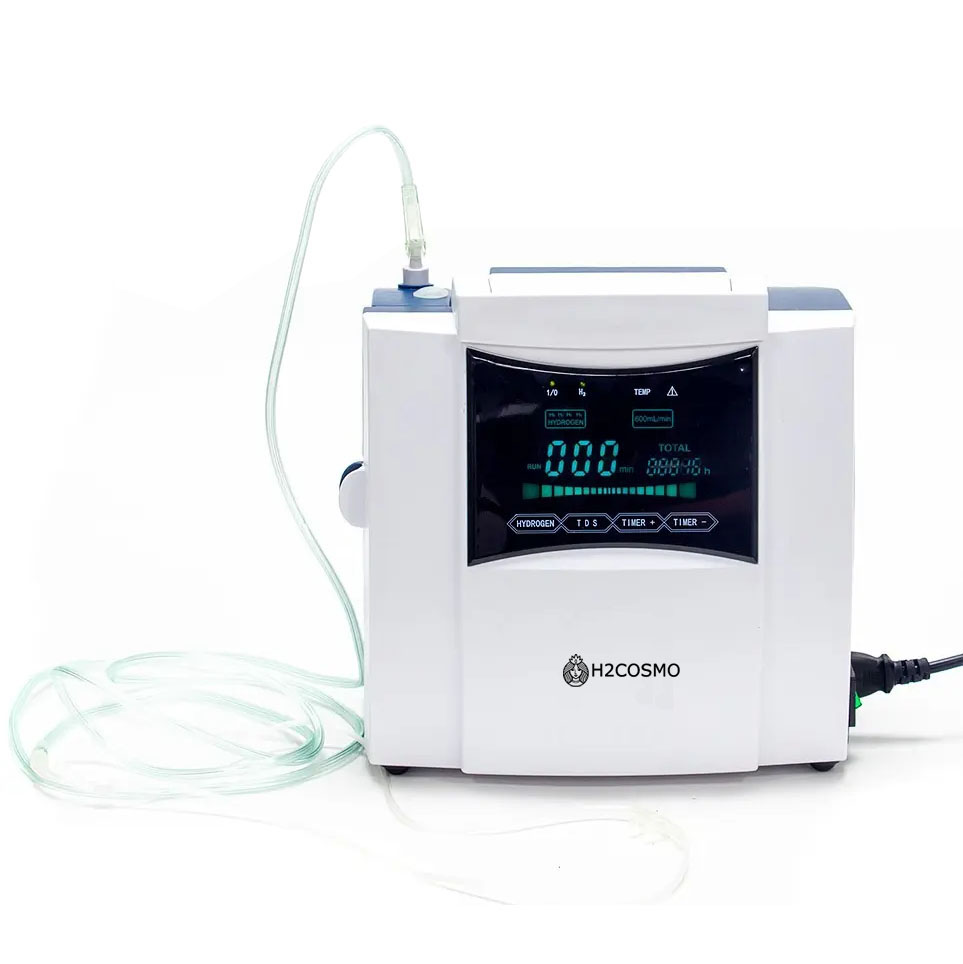
Scopri il potere rivitalizzante dell’acqua all’idrogeno con il nostro dispositivo all’avanguardia. Ogni sorso è un passo verso un benessere duraturo. Idratazione avanzata, antiossidanti più efficaci: trasforma la tua acqua quotidiana in una fonte di salute e energia!
Unisciti alla rivoluzione dell’idrogeno! Il nostro dispositivo utilizza la tecnologia più avanzata per infondere ogni goccia d’acqua con idrogeno puro, offrendoti non solo idratazione, ma anche benefici antiossidanti che possono aiutare a combattere l’invecchiamento cellulare e migliorare la tua vitalità generale.
Idrogeno Molecolare
L’acqua arricchita di idrogeno molecolare (H2) è una forma d’acqua in cui molecole di idrogeno gassoso sono disciolte in acqua. Questo tipo di acqua è divenuto popolare in alcuni ambienti di salute e benessere per i suoi benefici antiossidanti.
Idrogeno = Vita
In sintesi, l’acqua arricchita di idrogeno è un interessante esempio di come monro casino la ricerca scientifica possa portare allo sviluppo di nuovi prodotti per la salute e il benessere.
- Antiossidante: L'idrogeno molecolare (H2) è un piccolo atomo e un potente antiossidante. È capace di diffondersi rapidamente nei tessuti e attraversare le membrane cellulari per neutralizzare i radicali liberi, in particolare il radicale ossidrile, uno dei radicali liberi più dannosi e reattivi.
- Potenziali Effetti Salutari: Studi preliminari suggeriscono che l'acqua arricchita di H2 potrebbe avere effetti benefici, come la riduzione dello stress ossidativo, miglioramento della performance sportiva, diminuzione dell'infiammazione e protezione contro alcune malattie neurodegenerative. Tuttavia, è importante sottolineare che molte di queste ricerche sono ancora nelle fasi iniziali.


Transformazione

Rughe
L'idrogeno, essendo un antiossidante potente, offre benefici significativi per la pelle, tra cui la riduzione dell'invecchiamento cutaneo.


Acne
L'idrogeno molecolare aiuta a ridurre l'infiammazione nella pelle, che è un fattore chiave nella formazione dell'acne. L'acne è spesso aggravata da uno stress ossidativo eccessivo che danneggia le cellule della pelle e può peggiorare l'infiammazione. L'idrogeno, in qualità di antiossidante, aiuta a neutralizzare i radicali liberi e a ridurre il livello di stress ossidativo nella pelle.


Skin Care
L'idrogeno è in grado di neutralizzare i radicali liberi, particolarmente i radicali ossidrili, che sono tra i più dannosi per le cellule della pelle.



Antietà
L'idrogeno penetra facilmente nelle membrane cellulari per proteggere le cellule dai danni ossidativi, contribuendo così a mantenere l'aspetto giovanile della pelle.

Applicazioni Neuroprotettive
Alcuni studi suggeriscono che l'idrogeno potrebbe avere effetti protettivi sul cervello, potenzialmente utile in condizioni come l'ictus ischemico, il morbo di Alzheimer e il morbo di Parkinson.

Recupero da Affaticamento e Malattie
L'inalazione di idrogeno aiuta a ridurre l'affaticamento e accelerare il recupero dopo l'esercizio fisico o durante il recupero da malattie influenzali o raffreddori, migliorando l'efficienza energetica delle cellule.
Il Mondo H2Cosmo



Ero curiosa riguardo ai benefici dell'acqua all'idrogeno, quindi ho provato la macchina H2Cosmo. Devo dire che i risultati sono impressionanti! La mia pelle appare più idratata e ho notato una riduzione dell'infiammazione dopo i miei allenamenti. Il design della macchina è elegante e si adatta perfettamente al mio piano di lavoro in cucina. È un ottimo argomento di conversazione e adoro spiegarne i benefici ai miei amici e lettori!
Come allenatore di fitness, sono sempre alla ricerca di prodotti che possano supportare il recupero e la salute generale. La macchina all'idrogeno H2Cosmo è diventata un elemento fisso nella nostra palestra. I nostri clienti adorano il rapido aumento di idratazione che offre dopo sessioni intense. L'abbiamo da tre mesi ora, e la sua durabilità e prestazioni sono state eccellenti. È un investimento, ma i benefici per la salute valgono la pena.
TUTTE LE NEWS
Svago aristocratico nel Rinascimento italiano
Роль данных при совершенствовании пользовательского опыта взаимодействия
Intrattenimento signorile nel Rinascimento italiano
Proprietà antiossidanti e anti-infiammatorie
L’acqua idrogenata serve come un antiossidante, aiutando in varie malattie e riducendo le risposte infiammatorie (VerdeGusto).
Effetti sul metabolismo e la perdita di peso:
Bere acqua idrogenata innesca un meccanismo dimagrante chiamato termogenesi, aumentando il tasso metabolico e aiutando così a bruciare più calorie
Miglioramento della salute della pelle
Bere regolarmente acqua idrogenata potrebbe migliorare la salute della pelle, riducendo rughe, infiammazioni e rossore, e potenzialmente aumentando la produzione di collagene.
Riduzione del rischio di malattie croniche e cancro
L’acqua idrogenata ha mostrato di poter avere effetti benefici su malattie croniche e cancro, proteggendo il DNA e i mitocondri dal danno ossidativo
Benefici per reni e fegato
Bere regolarmente acqua idrogenata ha mostrato miglioramenti in pazienti con problemi epatici e renali.
Effetti Collaterali
Non ci siano effetti collaterali noti legati al consumo di acqua idrogenata o per inalazione, e l’idrogeno è generalmente ben tollerato dal corpo. Tuttavia, è importante ricordare che molte di queste affermazioni sono basate su studi preliminari e che ulteriori ricerche sono necessarie per confermare questi benefici in modo definitivo.
La maggior parte degli studi suggerisce il consumo di 0,5 – 2 litri di acqua idrogenata al giorno.
Descubre una amplia gama de juegos de casino y proveedores en topacio casino, donde puedes disfrutar de emocionantes tragamonedas, mesas con crupieres en vivo y atractivos bonos p.
Immergiti nel mondo dei rulli e delle vincite sorprendenti, dove i siti non aams ti aspettano con slot irresistibili e jackpot che potrebbero cambiarti la vita in un istante.
Scopri la varietà di giochi da casinò offerta da numerosi provider su https://sportitalia-bet.net/app/, e goditi l'emozione del gioco d'azzardo con le migliori slot e croupier dal.
Scopri un mondo di emozioni con una vasta gamma di giochi da tavolo e slot di prim'ordine su amazingbet casino, dove i migliori provider ti garantiscono un'esperienza unica e coin.
Scopri l'adrenalina del gioco online con incredibili slot e jackpot mozzafiato; su Smokace Italia, il brivido di vincere è a portata di mano con offerte uniche e dealer dal vivo!
Vivi l'emozione di vincere grandi premi giocando alle slot su Lizaro casino, dove i jackpot ti aspettano con sorprese mozzafiato e l'adrenalina è garantita ad ogni spin.
Immergiti nel brivido delle slot, dove i casinò online non aams offrono un'esperienza unica con jackpot incredibili e bonus irresistibili per i veri amanti del gioco d'azzardo.
Immergiti nell'atmosfera elettrizzante e autentica del sisal casino, dove i croupier dal vivo ti accompagnano in tempo reale tra slot emozionanti e tavoli da gioco coinvolgenti.
Registrati facilmente e scopri un'ampia gamma di giochi su Betscore casino, dove i depositi sono rapidi e i prelievi veloci garantiscono un'esperienza di gioco senza interruzioni.
Play your favorite casino games on the go with your smartphone or tablet, and enjoy the convenience of richard casino no deposit bonuses, making gaming both easy and rewarding.
Dopo aver usato la macchina all'idrogeno H2Cosmo per un mese, ho notato un notevole aumento dei miei livelli di energia. È stata un'aggiunta fantastica alla mia routine mattutina, aiutandomi a sentirmi rivitalizzato e pronto per la giornata. L'installazione è stata semplice e trovo che l'operazione della macchina sia incredibilmente facile da usare. La consiglio vivamente a chiunque voglia migliorare la propria routine di benessere!